The Neuromuscular Neutral Zone, Dynamic Stability, and Injury/Pain in the Course of Everyday Activity
Malik Slosberg, DC, MS
Understanding the dysfunctions that may destabilize joints offers new insights into the mechanisms that can be influenced by chiropractic adjustments to enhance dynamic joint stability. Recent research has brought into focus the critical relationship between unconscious sensory input from deep articular and periarticular structures and reflexive motor responses, which are essential to protect joints from injury. Let's review these new advances in our understanding of the neuromotor mechanisms that may go awry, making our patients vulnerable to injury and recurrence.
Ranges of Articular Motion Both Safe and Dangerous
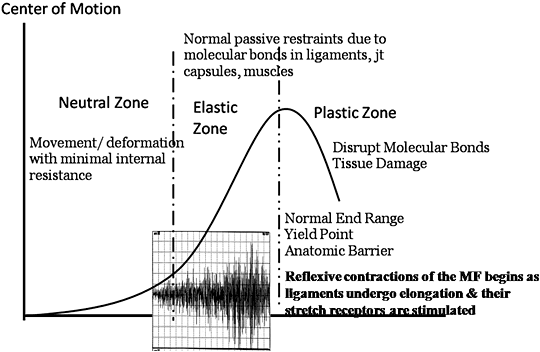
We are all aware of the basics of articular motion and the "zones" of motion around the center of joint motion or the joint's neutral position. However, are you familiar with the neuromuscular neutral zone? In the very recent literature understanding the neuromuscular neutral zone is implicated as a key factor in protecting and maintaining the integrity of spinal joints. As a review of the basic ranges of motion of a joint and their relationship to the neuromuscular neutral zone, let's begin with the neutral zone, the motion immediately around the mid-position of a joint.
The neutral zone is commonly defined as the range of displacement near the spine segments' neutral position, where minimal resistance is offered by the osteoligamentous structures.1 The authors of this definition caution that the neutral zone may increase with injury, articular degeneration, loss of passive stiffness, and weakness or inhibition of the stabilizing muscles. They warn us that when the neutral zone is increased the spine may become unstable and painful.
The next zone of motion is the elastic zone, defined as the zone that extends beyond the neutral zone within which tissues undergo increasing levels of physiological strain, but still remain less than sufficient to produce disruption or injury.2 As this strain increases, there is progressive internal resistance to further deformation as a result of the molecular bonds in the connective tissue matrix of ligaments, joint capsules, intervertebral discs and muscles in order to prevent injury. The structures that resist possible injurious deformation have been referred to as the "normal restrictive " or "physiologic" barriers."3
The molecular bonds within the connective tissue matrix protect joints from becoming over-distracted and injured. If the forces applied are too great for these molecular bonds to tolerate, the joint is forced beyond the limits of its normal range of motion into the "plastic zone," in which disruption of molecular bonds and tissue injury occurs; the higher the forces and the further beyond the normal end-range of motion a joint is forced, the greater the amount of tissue disruption and injury.
Disruption of Molecular Bonds and Joint Injury
Typically, the molecular bonds first disrupted are those within the passive restraints of the joint; that is, the ligaments, joint capsules, and intervertebral discs. These structures resist the tensile or torsional forces a joint is subjected to and which may cause damage if not well-controlled. The initial injuries documented to occur if the joint is forced beyond its normal end range have been variously labeled as "subfailure injury,"4 "microdamage of collagen tissues"5 and "microruptures of collagen fibers."6 These collagenous matrices provide the passive restraints that attempt to protect joints from being forced into the plastic zone.
Dynamic Stability and Protective Reflexive Motor Responses
The research is clear that it is important for the active restraints - dynamic muscle contraction - to be rapidly and reflexively elicited in order to protect the passive restraints from damaging overload when a sudden or unexpected load is applied. Appropriately timed muscle contraction can dampen and absorb some of the load or stress to which the ligaments are subjected and, by doing so, reduce the risk of damage to collagen fibers.
Reflex contractions of the back muscles, especially of the multifidus, normally act to control spinal movements in order to prevent excessive loading and to protect the underlying tissues from injury.7 The sensory-motor interactions between ligament tension and muscle recruitment occur very rapidly and unconsciously in order to ensure safe and controlled movement. Terminology referring to this interaction includes the ligamentomuscular reflex or facet joint capsule muscular reflex.8
Neuromuscular Neutral Zone and Triggering of the Multifidus
The neuromuscular neutral zone is defined by the tension or displacement in the lumbar spine at the instant that reflexive EMG activity is first triggered in the multifidus during flexion or stretch.5 The question arises: What precisely triggers this reflexive motor activity? With deformation, passive ligamentous restraints become stretched and, as they do, stretch receptors within them become stimulated. Impulses initiated from these receptors instigate a protective spinal reflexive motor response by the extensor muscles, particularly the multifidus, the first muscle to contract and protect the passive restraints from injury.9 The active and passive stabilizers are interactive due to their sensory-motor feedback loop, a synergystic relationship to provide dynamic load sharing under various conditions.5
According to recent research, the multifidus accounts for two-thirds of the control of lumbar intervertebral motion.10 In addition, the multifidus, because it is segmentally innervated, has the capacity for fine control of movement of individual lumbar vertebrae. Each nerve innervates only the fascicles that arise from that vertebrae, indicating a direct relationship between a segment and its multifidus. The segmental multifidus can, therefore, control a segment to match the applied load.11
Difficulties arise when this reflexive motor response is delayed and begins too late to adequately dampen the tensile and/or torsional forces. As a result, molecular bonds within the connective tissue matrix of ligaments, joint capsules and/or discs are strained beyond their elastic limits and disrupted; a sprain occurs. According to evidence on the mechanisms of ligaments sprains, these subfailure injuries account for more than 85 percent of all ligament injuries.12
Why Is the Protective Reflexive Motor Response Delayed?
There are several factors that come into play and help explain the delay. One primary mechanism, which has been extensively documented, has to do with repetitive movements, particularly in sagittal flexion and/or flexion in combination with rotation. With repetitive activity, ligaments undergo creep deformation/ligament laxity, which results in changes in responsiveness of the mechanoreceptors within the ligaments leading to a "false kinesthetic perception,"6 thereby delaying protective reflexive motor responses. As Solomonow, et al.,8 described in their Volvo Award-winning article:
"Laxity in the ligaments desensitizes the mechanoreceptors within them and, therefore, significantly diminishes or completely eliminates their ability to monitor relative intervertebral motion, rendering them unable to reflexively initiate the muscular forces within the multifidus necessary to prevent destabilizing motion. Once laxity in the ligaments is present, the spine does not benefit from any protective muscular activity, and is exposed to injury."
Creep Deformation: How It Affects Sensory and Motor Responses
A 2010 paper7 describes the changes in ligamentous tissues resulting in creep as a time-dependent expulsion of water from spinal tissues, especially intervertebral discs, causing loss of disc height, slack in posterior ligaments and reduced resistance to bending by the osteoligamentous spine. With increased laxity, passive tissues do not elicit the same neural response as when tissues are taut. Stretch receptors in the posterior ligaments become stimulated when ligaments are stretched and the impulses initiate a spinal reflexive motor response by the extensor muscles. The decreased activity of the stretch receptors decreases activity of the muscles and together with laxity in the ligament, leads to increased laxity at the joint.9
Soft-tissue creep causes delayed activation of the back muscles in response to sudden loading. Ligament laxity alters afferent feedback from ligament receptors and impairs reflex activation of back muscles. Impaired reflexes in the back muscles as a result of prolonged or repeated flexion could increase the risk of bending injuries to the spine.7
Clinical Relevance
A practical application of this avenue of research is this: One of the most common presentations of low back pain results from the uneventful engagement in prolonged activity over the course of a day, yet after the repetitive tasks, work or athletic/exercise activity is over, people describe that while they are changing clothes, twisting to get into or out of their car, bending over to tie their shoes, or performing other everyday movements, they sense a distinct "pop" in the low back followed by pain. Note that this onset of pain is not associated with any external load, slip or fall, or unexpected application of force. The microdamage to collagen fibers occurs not as the result of excess, rapidly applied, or unexpected external load, but is due to poor internal neuromotor control of the upper-body torso mass under dynamic conditions.
This research confirms that a significant deficit in passive and active stability is present for hours immediately after a prolonged work period or repetitive flexion. Some researchers conclude that it is not surprising or may even be expected that minor or routine movements may result in instability and injury.7 From this information, clinicians can better understand what and why tissue damage may occur and they can better manage these patients and promote their full recovery, including restoration of functional capacity. This will be the focus of my next column.
References
Kolber MJ, Beekhuizen K. Lumbar stabilization: an evidence-based approach for the athlete with low back pain. Strength and Conditioning Journal, 2007;29:26-37.
McDonald D, Moseley GL, Hodges PW. Why do some patients keep hurting their back? Evidence of ongoing back muscle dysfunction during remission from recurrent back pain. Pain, 2009;142:183-188.
Richardson, Hodges, et al. Therapeutic Exercise for Lumbopelvic Stabilization. A Motor Control Approach for the Tx & Prevention of LBP. Edinburgh, Churchill Livingstone, 2004:59-73.
Building on a historic March 2026 meeting between Make America Healthy Again and chiropractic leadership, MAHA has announced the launch of the MAHA Chiropractic Hub, “a coordinated national partnership uniting MAHA Center, MAHA Action, and the chiropractic profession, including national associations, state organizations, practitioners, educators, researchers, and patient advocates. The Chiropractic Hub will advance federal policy, expand patient access, and build broad public support for chiropractic care across America.”
The chiropractic profession is confronting one of the most significant federal regulatory threats it has faced in decades. A new U.S. Department of Education (ED) accountability framework – commonly referred to as the “Do No Harm” (DNH) regulation – could place many chiropractic programs at risk of losing access to federal financial aid (student loans), potentially reshaping the future of chiropractic education and workforce development across the United States.
Pain has become the dominant language of musculoskeletal healthcare. Numeric pain-rating scales and symptom reports are routinely used as primary indicators of clinical success. But while pain reduction is meaningful, it is an incomplete and often misleading representation of recovery. This has real consequences for patient adherence, long-term outcomes, and how conservative care is perceived within the broader healthcare system.